キク科Flaveria属の分子系統樹の完成
C₄光合成進化に関わる新たな仮説の提唱
キク科Flaveria属の分子系統樹の完成
C4光合成進化に関わる新たな仮説の提唱
国立大学法人東京農工大学大学院農学研究院生物生産科学部門 安達俊輔准教授と同グローバルイノベーション研究院スーパー教授(カナダトロント大学教授)Rowan F. Sageらは、高効率な光合成(C4光合成)進化のモデル植物であるキク科Flaveria属植物を網羅する分子系統樹を作成しました。さらにそれら植物の光合成特性や酵素特性の解析を通じて植物がC4光合成を獲得した過程に関わる新たな仮説を提唱しました。C4光合成をもたないイネに人為的にC4光合成を導入する試みが世界中で行われており、本研究成果はその実現の後押しとなるものです。
本研究成果は、Plant Physiology誌に10月6日に掲載されました。
論文名:The Evolution of C4 Photosynthesis in Flaveria (Asteraceae): Insights from the Flaveria linearis Complex
著者:Shunsuke Adachi, Matt Stata, Duncan G. Martin, Shifeng Cheng, Hongbing Liu, Xin-Guang Zhu and Rowan F. Sage
掲載URL:https://academic.oup.com/plphys/advance-article/doi/10.1093/plphys/kiac467/6749591
現状
トウモロコシやサトウキビなどのC4植物
注1)
は、葉肉細胞と維管束鞘細胞が協働する独自の光合成システム(C4光合成)を有しており、葉肉細胞のみで光合成を行う(C3光合成)イネ、コムギ、ダイズなどのC3植物と比較して高い光合成効率を発揮します。そのため、C3植物である作物にC4光合成を導入することによって、生産性の大幅な向上が期待されています。地球上に現存するすべてのC4植物はC3植物から進化したと言われており、しかも多様な植物系統において60回以上独立に進化(いわゆる収斂進化
注2)
)したことが知られています。この進化メカニズムの詳細を解明することが、イネなどの作物に対する人為的C4化に貢献すると考えられています。
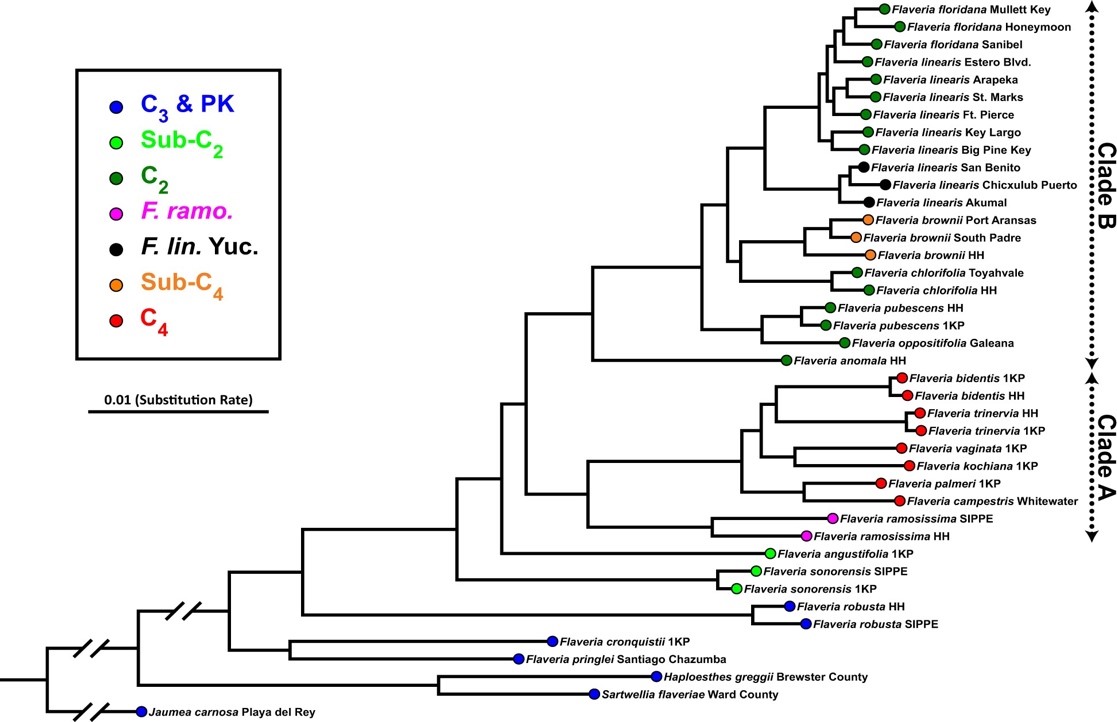
キク科の植物であるFlaveria属には、C3植物からC4植物への光合成進化の痕跡を残す種が数多く含まれています。たとえばC3植物のF. pringlei, C2植物 (C3-C4中間植物) のF. linearis、C4-like植物 (C4に非常に近い) のF. brownii、C4植物のF. trinerviaなど計21種が知られています (図1)。Flaveria属はC4光合成進化のモデル植物として長年にわたり研究対象となってきました。しかし、過去の研究ではこれらの植物種の一部を利用したものが多く、網羅的な分子系統樹
注3)
はありませんでした。
研究体制
本研究はUniversity of Toronto, Faculty of Arts & Science, Department of Ecology & Evolutionary Biology, Rowan F. Sage研究室および東京農工大学大学院農学研究院生物生産科学部門で実施しました。
研究成果
世界中から収集したFlaveria属の植物コレクション(Flaveria属の種や亜種、近縁種を含む全45系統)をトロント大学の温室内で栽培し、実験を行いました。まずゲノムデータに基づき網羅的なFlaveria属の分子系統樹を完成させました(図2)。従来の報告と同様、Flaveria属の種が2つの大きな分類群に分けられることが確かめられました。一方で、F. browniiの最も近縁な種はF. chlorifoliaであること、メキシコのユカタン半島に自生するF. linearisはフロリダに自生するF. linearisとは別種である可能性が高いこと、F. floridanaはフロリダに自生するF. linearisと同種である可能性が高いことなど、従来の説を覆す発見がありました。
これらの植物のC4光合成酵素の活性を、分光光度計を用いた方法により測定しました。ホスホエノールピルビン酸カルボキシラーゼやNADP-リンゴ酸酵素などの活性はC4-like植物やC4植物において極めて高く、C3植物やC3-C4中間植物とは大きな開きがありました。これは、C3-C4中間植物がC4植物に進化する過程において、C4酵素活性のダイナミックな変化があったことを示しています。一方、これまでC4光合成システムの脇役と捉えられていたアスパラギン酸アミノ基転移酵素やアラニンアミノ基転移酵素の活性は、C3植物からC3-C4中間植物、C4植物にかけて連続的に上昇していることがわかりました。筆者らは、これらの酵素活性の高まりがホエノールピルビン酸カルボキシラーゼの基質であるピルビン酸の再生を促し、C4光合成の形成に結びついたとする新たな仮説を提唱しました。
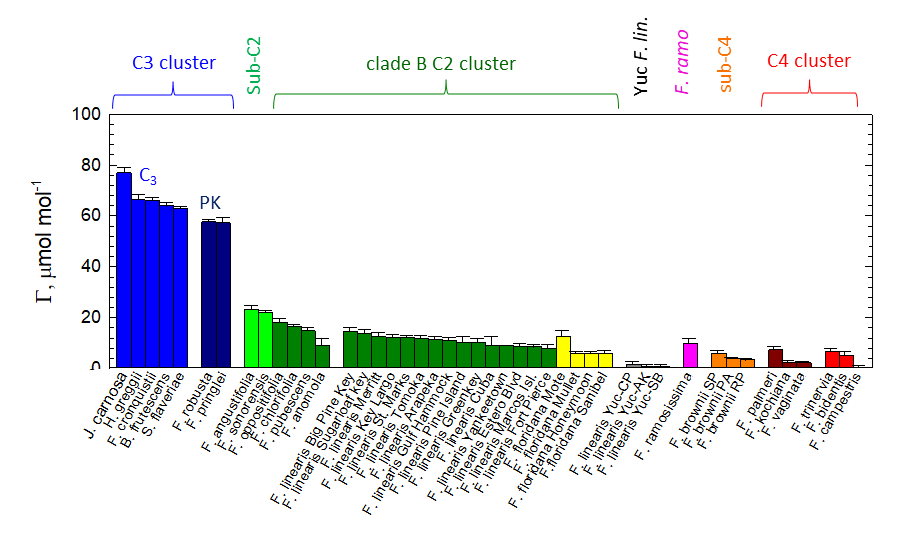
葉のCO2ガス交換測定(光合成測定)結果から、もう一つ興味深い結果を得ました。メキシコのユカタン半島に自生するF. linearisは、フロリダに自生するF. linearisと比較して極めて小さなCO2補償点
注4)
を持ち、見かけ上光呼吸
注5)
を生じていませんでした (図3)。光呼吸とはC4植物を除くすべての植物において光合成の副産物として生じる代謝反応で、光合成効率を低下させることが知られています。すなわちC3-C4中間種である本種は、光呼吸を抑制する独自のシステムを発達させてきたことになります。ユカタン半島の海岸沿いという高温や塩ストレスを受けやすい地域において、適応度を高めるため独自の進化を遂げたものと考えられます。
今後の展開
C3植物からC4植物への進化過程は、植物科学における最大の謎の一つです。今回の研究で提唱された仮説が検証されれば、C4光合成進化メカニズムやそれをもたらした鍵遺伝子が解明に結びつき、イネなどの主要作物のC4化の実現に一歩近づくと期待されます。またユカタン半島に自生するF. linearisの光呼吸抑制システムが解明されれば、光合成進化の新たな過程が明らかになるかもしれません。
用語解説
注1) C4植物 トウモロコシ、ソルガム、サトウキビ、ヒエなど、C3植物とは異なる独自の光合成システム (C4光合成) を持つ植物の総称。
注2) 収斂進化 種の系統が異なっても、環境に適応して類似の外見や機能を有するようになること。
注3) 分子系統樹 生物の持つDNAなどを比較することで生物間の系統関係を示した樹形図のこと。
注4) CO2補償点 光合成によるCO2吸収速度と光呼吸・暗呼吸によるCO2放出速度が釣り合うときのCO2濃度のこと。この値が小さいほど呼吸 (主に光呼吸) によるCO2ロスが小さい。
注5) 光呼吸 CO2固定酵素であるRubiscoが持つO2固定活性により産生された毒性物質(ホスホグリコール酸)を除去するために働く一連の化学反応のこと。反応過程で細胞内からCO2が放出されるためにこの名が付いた。
研究費
本研究はグローバルイノベーション研究院からの支援ならびにJSPS科学研究費補助金(16K18643 および 21K19104)の支援を受けて行われたものです。

◆研究に関する問い合わせ◆
東京農工大学大学院農学研究院
生物生産科学部門 准教授 安達 俊輔(あだち しゅんすけ)
TEL:042-367-5671
E-mail:adachi(ここに@を入れてください)go.tuat.ac.jp