大発生のタイミングはお見通し?~ハバチの不規則な発生に同調する寄生バチを発見~
大発生のタイミングはお見通し?
~ハバチの不規則な発生に同調する寄生バチを発見~
ポイント
- ハバチの不規則な発生に同調して発生する寄生バチを発見した。
- 寄生バチのハバチへの寄生率は、ハバチの密度の違いにかかわらず一定であった。
- 寄生バチは自身の発生の程度をハバチの発生の程度に同調させることで、長期的に安定してハバチに寄生しているとみられる。
本研究成果は、アメリカの昆虫学誌「Biological Control(略称:Biol. Control)」オンライン版(12月1日付)に掲載されました。
論文名:Response of specialist and generalist predators to nonprogressive annual fluctuations in herbivorous insect populations.
著者名:Toru Taniwaki, Kyohei Watanabe, Hirotaka Komine, Kahoko Tochigi, Masanobu Yamane, Shinsuke Koike
URL:https://doi.org/10.1016/j.biocontrol.2021.104810
概要
国立大学法人東京農工大学大学院農学府の谷脇徹産学官連携研究員(当時、現在は神奈川県自然環境保全センター主任研究員)、同大学院グローバルイノベーション研究院の小池伸介教授、同研究院の小峰浩隆特任助教(当時、現在は山形大学)、神奈川県立生命の星・地球博物館の渡辺恭平主任学芸員、神奈川県自然環境保全センターの山根正伸主任専門員らの共同研究チームは、ハバチとその天敵である寄生バチの生態を調べ、ハバチは年ごとに不規則に発生しているにもかかわらず、ハバチの発生の程度に同調して発生する寄生バチがいることを初めて発見しました。この寄生バチのハバチへの寄生率はハバチの発生の程度にかかわらず、おおむね一定を維持していました。寄生バチは不規則に発生するハバチの発生の程度を何らかの方法で察知し、自身の発生する程度を同調させることで、長期的に安定してハバチを宿主(注1)として利用していると考えられます。
研究背景
森林では植食性昆虫(注2)が数年に一度のタイミングで大発生し、樹木の葉が食べつくされることがあります。植食性昆虫のなかでも、ハバチ(葉蜂)類(注3)は不規則な間隔で、突発的に大発生することが知られていますが、どのような要因により大発生が生じているのかは不明でした。このように謎の多いハバチ類の大発生の要因を解明するには、まずはハバチ類の主要な死亡要因である天敵となる昆虫(以下、天敵昆虫)(注4)が、ハバチの大発生時にどのように発生の程度を反応させているのかを解明する必要があります。天敵昆虫の中には、特定の種類の昆虫を餌として利用する傾向の強いスペシャリスト(注5)と、様々な種類の昆虫を幅広く餌として利用するジェネラリスト(注6)が存在します。一般的に、天敵昆虫は餌となる昆虫の生息数が増えると、一定の時間が経った後に、同じように生息数が増えるとされてきました。しかし、突発的に大発生するハバチ類に対して、2つのタイプの天敵昆虫がどのように生息数を変化させるのかは分かっていませんでした。

研究体制
本研究は丹沢山地のブナハバチ対策に取り組む神奈川県自然環境保全センターと東京農工大学、神奈川県立生命の星・地球博物館の共同研究として行われました。
研究方法
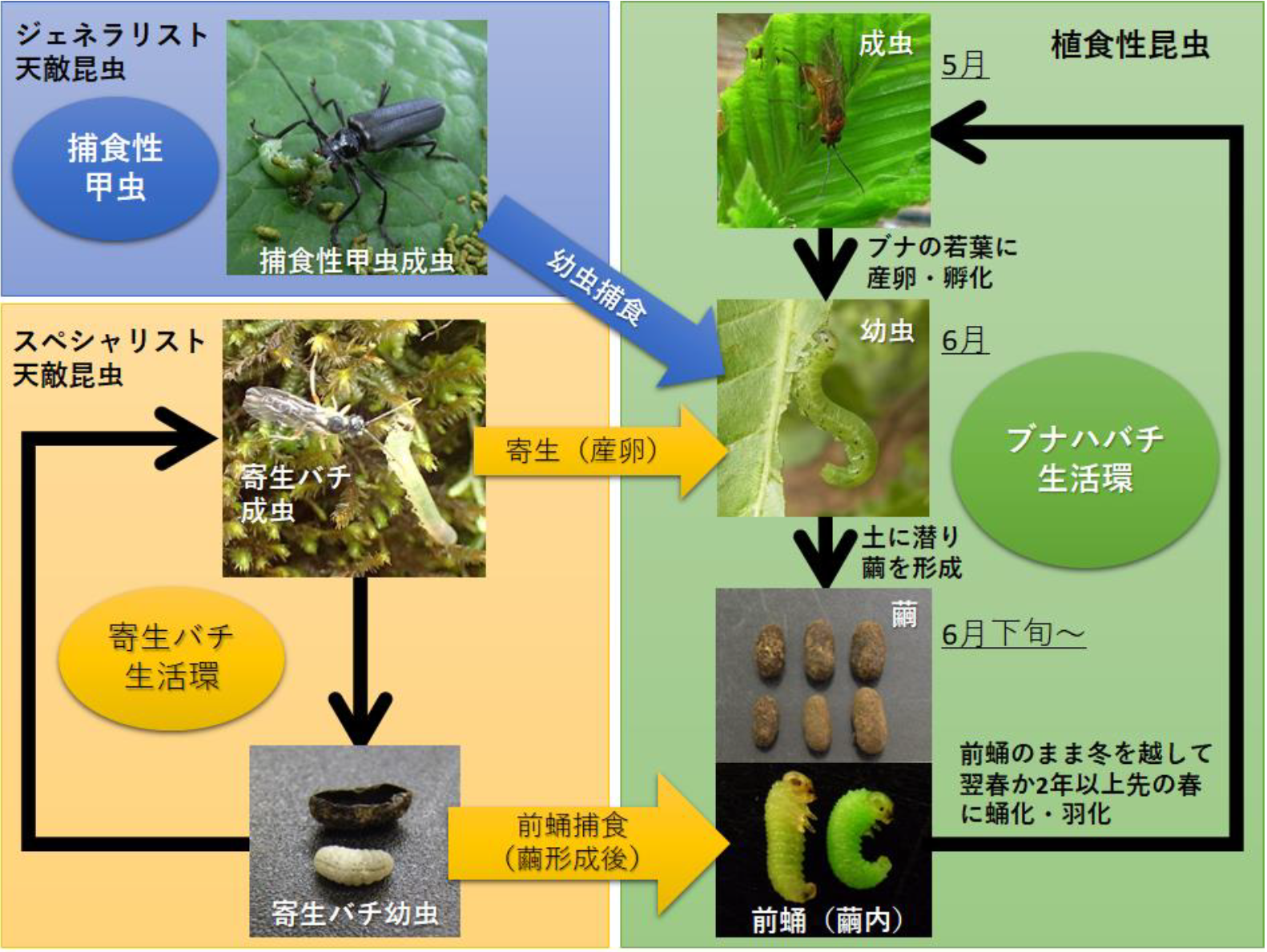
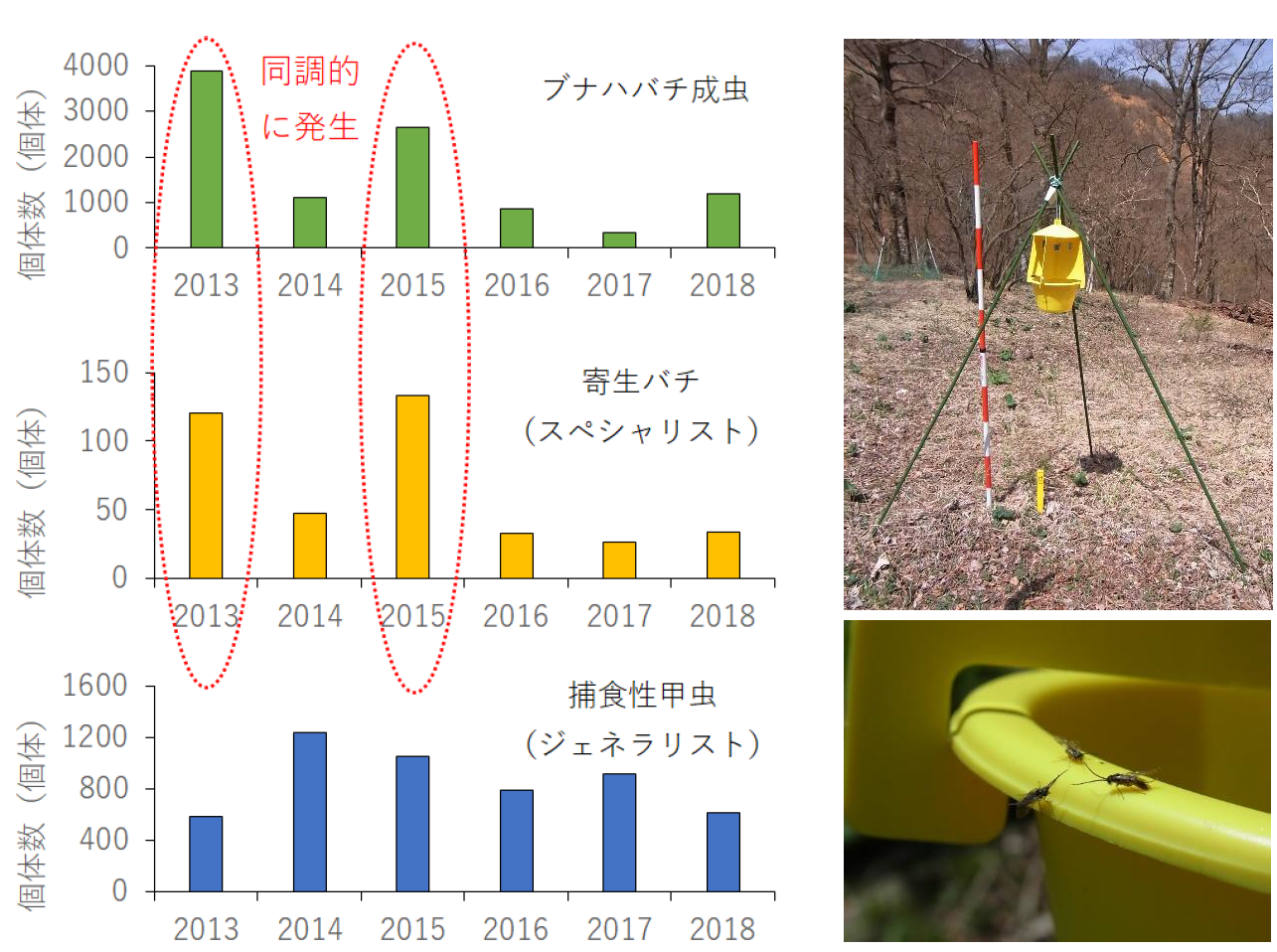
ハバチの大発生自体は自然の現象ですが、神奈川県の丹沢山地では大気汚染などで衰弱したブナ林において、ブナハバチというハバチの1種(図1、2)が大発生することで、ブナの枯死が促進されることが問題となっていました。そこで、丹沢山地のブナ林におけるブナハバチに着目し、その天敵昆虫のうちスペシャリストである寄生バチ(キモンブナハバチヤドリヒメバチなどのヒメバチ類)(図2)と、ジェネラリストである捕食性甲虫(クロジョウカイなどのジョウカイボン類)(注7)(図2)の、ブナハバチの大発生時およびその前後の生息数の変化を調べることにしました。具体的には、昆虫捕獲器(注8)(図3)を用いて、2013年から2018年にかけての4月下旬から7月上旬に、ブナハバチ、寄生バチおよび捕食性甲虫を採取するとともに、2011年から2018年にかけての秋季にはブナハバチが土中で形成した繭の数と、それらの繭に対する寄生バチによる寄生の割合を調べました。
ブナハバチは丹沢山地では主に5月に成虫となり、その後産卵された卵は6月には幼虫となります(図2)。捕食性甲虫はこの幼虫を捕食し、寄生バチはこの幼虫に寄生(産卵)します(図2)。捕食されたブナハバチの幼虫はそのまま死亡します。一方、寄生(産卵)されたブナハバチの幼虫はすぐには死亡しません。その後、ブナハバチの幼虫は土に潜って繭を形成し、蛹の前の段階である前蛹(ぜんよう)という発育段階で過ごしますが、繭の中におけるこの発育段階において寄生バチの幼虫に捕食されて死亡します(図2)。その後、寄生バチの幼虫はブナハバチの繭の中で、新たに自身の繭を形成します。
研究成果
ハバチと寄生バチの同調的な発生
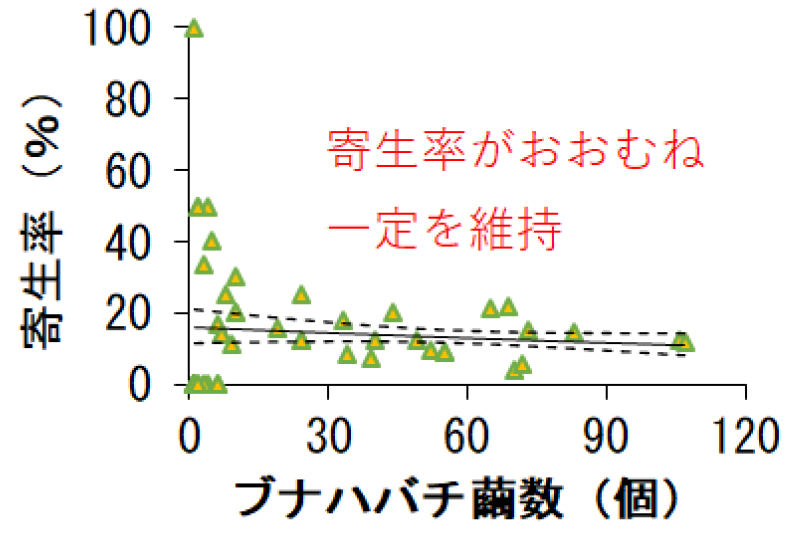
これまで植食性昆虫の一つであるガ類では、その生息数が増えると天敵昆虫も一定の時間が経過した後に、同じように生息数が増えることが知られていました。そこで当初は、ブナハバチの2つのタイプの天敵昆虫もまた、不規則に大発生するブナハバチに対して、一定の時間が経過した後に、同じように大発生すると考えていました。実際、ジェネラリストである捕食性甲虫の生息数は、反応の程度は弱いながらも、ブナハバチの生息数が増えると、遅れて生息数が増えることが認められました(図3)。ところが、スペシャリストである寄生バチは、ブナハバチの発生に遅れることなく、同じ年に同じような程度で発生数が変化していました(図3)。さらに、ブナハバチの繭の寄生バチによって寄生された割合は、年や場所によってブナハバチの生息数(繭の数)が変化しても、おおむね一定の割合を維持していました(図4)。このことは、寄生バチがその発生の程度をブナハバチの発生と同調させることで、ブナハバチを長期にわたり、安定的に寄主として利用していることを強く示し、世界でも前例のない植食性昆虫と天敵昆虫との種間関係の発見となりました。
ハバチと寄生バチにみられる長期休眠の生態
それでは、なぜこのような植食性昆虫とスペシャリスト天敵昆虫の間で、発生数の変化の同調が生じているのでしょうか。ヒントは長期休眠(注9)の生態にあると考えています。
ブナハバチは卵から成虫に至る1世代に要する期間が、個体により1年~4年と変化します。ブナハバチは初夏から春にかけて、土中の繭の中で前蛹の状態で過ごしますが、成虫になるまで2年以上の時間を要する個体では、春になっても蛹にはならず、前蛹のまま翌年の春、あるいは翌々年の春まで休眠を継続します(図2)。このように休眠の期間が個体によって異なることで、産卵の翌春に成虫になる個体の割合が年によって変化し、その結果ブナハバチにも不規則な大発生が生じている可能性が考えられます。
さらに、ブナハバチの繭の中でブナハバチの前蛹を捕食した寄生バチの幼虫の中にも、寄生した翌春以降も自身が作った繭の中で休眠を継続する個体の存在が確認されました。以上から、ブナハバチと寄生バチの両者の発生の程度に同調が生じているメカニズムとして、ブナハバチを餌として利用する寄生バチが、ブナハバチの休眠期間や羽化のタイミングを何らかの方法で察知することで、自身の発生量の程度を調整している可能性があります。
長期休眠の長さに影響する要因
ブナハバチ、寄生バチとも、その休眠期間や成虫になるタイミングが、どのような要因によって決まるのかは、まだよく分かっていません。ただし、ブナハバチの大発生は丹沢山地の中でも広く同調するうえ、中には20~100km離れた場所でも、大発生が同調することが知られています。したがって、気象条件のような広範囲で同じように変化する要因が、ブナハバチの休眠期間や成虫になるタイミングに作用し、さらに寄生バチにも同じように作用している可能性があります。あるいは、寄生バチは、宿主であるブナハバチの何らかの生理的な状態の変化から、ブナハバチの休眠期間や成虫になるタイミングを察知しているのかもしれません。今後、これらの要因の検証や具体的なメカニズムを明らかにしていく必要があります。
今後の展開
本研究で得られた植食性昆虫―天敵関係の新たな知見は、昆虫の長期休眠の生態研究や、寄主である昆虫の天敵昆虫に対する適応といった、進化学的研究の発展に大きく寄与することが期待されます。また、ブナハバチの大発生に対して、自然に存在するいずれのタイプの天敵昆虫も、ブナハバチの大発生を抑えたり、その翌年にブナハバチを大きく減少させるような大発生をすることはないことが見えてきました。以上の知見から、ブナハバチの大発生の要因の解明が進むことが期待されます。さらに、丹沢山地のブナ林の保全の観点からは、様々な要因で衰退しているブナ林においては、ブナハバチの大発生時には、出来るだけ早めにその兆候をつかむとともに、環境に負荷のかからない方法でブナハバチの発生数を抑えるなどの対応策を考えていく必要があります。
用語説明
注1) ある生物がほかの生物の体内または体表を生息場所とし、おもに栄養面においてその生物に依存して生活することを寄生といい、寄生する側を寄生者、寄生される側を宿主という。
注2) 植物の葉や汁などを餌とする昆虫のこと。ハバチのほか、ガやチョウ、ハムシ、アブラムシなど様々な種類が知られている。
注3) 植物を食べる原始的なハチの仲間で、植物体に切れ込みを入れて産卵するための平たいノコギリ状の産卵管を持つ。動物を刺すための針は持たない。
注4) 昆虫やクモなどの節足動物を捕まえて食べる昆虫や、節足動物の体に寄生して養分を摂取し最終的にその寄主を殺してしまう昆虫のこと。前者ではトンボやカマキリ、後者では寄生バチや寄生バエなどが知られている。
注5) 特定の種類の昆虫しか餌として利用しない昆虫。一般的に、餌となる種類の昆虫の密度変化の影響を受けやすい。
注6) いろいろな種類の昆虫を餌として利用する昆虫。特定の種類の昆虫との結びつきが弱く、餌として利用する、ある種類の昆虫が低密度のときでも、他の種類の昆虫を餌として利用することで、その密度を維持することができる。
注7) 昆虫やクモなどの節足動物を捕まえて食べる昆虫の中でコウチュウ目に属する昆虫。ジョウカイボン類のほか、テントウムシやオサムシなどが知られている。
注8) 昆虫を捕獲する道具(トラップ)。屋根と十字に組み合わせた2枚の板とバケツで構成される、それらの黄色に誘引されたブナハバチ、寄生バチ、捕食性甲虫が板に衝突して落下し、バケツ内の溶液の中に落ちて、捕えられる。通称、衝突板トラップ。
注9) 昆虫は冬の低温などの厳しい季節をやり過ごすために休眠するが、昆虫の種類によっては、一部の個体が休眠状態で冬を過ごしたのち、春以降に成長を再開せずに休眠を継続することがある。そのような状況を長期休眠という。
◆研究に関する問い合わせ◆
東京農工大学大学院グローバルイノベーション研究院 教授
小池 伸介(こいけ しんすけ)
E-mail:koikes(ここに@を入れてください)cc.tuat.ac.jp
神奈川県自然環境保全センター研究企画部研究連携課 主任研究員
谷脇 徹(たにわき とおる)
E-mail :taniwaki.6y3j(ここに@を入れてください)pref.kanagawa.jp
関連リンク(別ウィンドウで開きます)
•東京農工大学 小池伸介教授 研究者プロフィール
•東京農工大学 小池伸介教授 研究室ウェブサイト
•小池伸介教授が所属する 東京農工大学農学部地域生態システム学科