〔2015年11月9日リリース〕体の左右を決める繊毛の構造解析とコンピューターシミュレーションにより繊毛が回転運動するしくみを解明
東京農工大学大学院工学研究院生命機能科学部門の篠原恭介特任准教授と大阪大学大学院生命機能研究科の濱田博司教授は、北京理工大学、大阪大学超高圧電子顕微鏡センター、理化学研究所ライフサイエンス技術基盤研究センターと共同で体の左右非対称性を決める繊毛の回転運動のメカニズムを明らかにしました。これまでに実験的な知見の少なかった繊毛構造と繊毛運動パターンの関係性の一端が明らかとなり、この成果により、今後、繊毛・鞭毛運動の不全が引き起こす内蔵逆位、気管支炎や不妊などの疾病の原因解明が期待されます。
|
本研究成果は、米科学誌Developmental Cell誌(10月26日付:日本時間10月27日)に掲載されました。 |
現状:われわれヒトに最も近いモデル動物であるマウスにおいて体の左右は受精後8日目に決定されます。この時期、体の正中線上に現れるノードと呼ばれる約200の細胞群がもつ繊毛(長さ2-5 mの運動性をもつ細胞から突き出た構造)が時計回りに回転し将来の体の右側から左側へ向かう水の流れ(ノード流)を発生させ、この流れがノードの左右で発現していた遺伝子の対称性を破ることで体の左右を決めると考えられています。この時期までにマウス胚は前後の極性が決まっていて、左向きのノード流は、(1)体の後ろ方向に傾いた回転軸と(2)時計回りの回転運動の2つを組み合わせる事で生み出しています。ノードの細胞の前後極性を決定するしくみについては近年PCP (Planar Cell Polarity:平面内細胞極性)シグナルの関与が報告されています。一方で繊毛が時計回りに回転運動するしくみについては不明でした。
研究体制:篠原恭介 (東京農工大学大学院工学研究院生命機能科学部門特任准教授・元大阪大学大学院生命機能研究科助教)、 Duanduan Chen (北京理工大学准教授)、 西田倫希 (一般財団法人日本繊維製品品質技術センター・元大阪大学超高圧電子顕微鏡センター特任研究員)、 美崎佳寿代 (理化学研究所ライフサイエンス技術基盤研究センター・テクニカルスタッフ)、 米村重信(理化学研究所ライフサイエンス技術基盤研究センター・チームリーダー)、 濱田博司(大阪大学大学院生命機能研究科教授)
研究成果:今回、抗がん剤であるタキソール(微小管重合に作用する)をマウス胚に処理するとノード繊毛の運動パターンが乱れるという現象を発見しました。繊毛の構造を大阪大学超高圧電子顕微鏡センターの保有する超高圧電子顕微鏡により調べた所、通常ノード繊毛の微小管は膜近傍に規則正しく配置されているのに対し、タキソール処理をしたノード繊毛では微小管の配置の規則性が乱れる事が分かりました。さらに構造と運動パターンの因果関係を明らかにするため、電子顕微鏡の実験データを元にした繊毛運動のコンピューターシミュレーションを行った所、微小管の配置が異常となると運動パターンも乱れる事が分かりました。この事からノード繊毛が安定に一方向に回転運動をするためには微小管の規則正しい配置が必要である事が分かりました(図1)。マウス運動繊毛は中心構造を持たないノード繊毛と中心対微小管・ラジアルスポークを持つ運動繊毛(気管・卵管・脳室など)の2種類の運動繊毛を持ちます。ノード繊毛がタキソール処理によって構造が変化してしまうのは、微小管配置を支える役割を担う中心構造を元々持たないためであるという仮説をたてその検証を行いました。まず野生型マウス気管繊毛にタキソール処理を行った所、繊毛運動や構造に変化は認められませんでした。次にラジアルスポークの頭部のタンパク質をコードする遺伝子を破壊したノックアウトマウス(Rsph4a
KOマウス)を作製しました。気管繊毛運動を観察した所、野生型マウスでは平面内に運動するのに対してノックアウトマウスの気管繊毛はノード繊毛と同様に時計回りに回転運動をしていました。最後にこのノックアウトマウスの気管繊毛にタキソールを処理したところ、繊毛の微小管に配置がタキソールに反応する事で乱れ、時計回りの回転運動が方向の定まらない回転運動に変化しました。これらの事から、ラジアルスポークが周辺微小管と中心構造を物理的につなぐ事により周辺微小管の規則正しい配置を安定に維持している事が示唆されました(図2)。ノード繊毛はマウス胚の前後極性の情報を元に左右極性を作り出す必要があります。そのためには平面運動ではなく回転運動が必須となるため、進化の中で中心構造をあえて失ったと考えています。
今後の展開:今回の成果によりノード繊毛の一方向に回転運動するしくみが明らかとなりましたが、なぜ回転方向が時計回りなのか、どのようにしてラジアルスポークは回転運動を平面運動へ変換しているのかといった基本的な疑問が残ります。今後これらの問題解明に着手する事で、哺乳類の運動繊毛の運動パターンを決める原理のより深い理解とともに、繊毛運動の不全が引き起こす内蔵逆位・気管支炎や不妊の疾病の原因解明につながると考えられます。
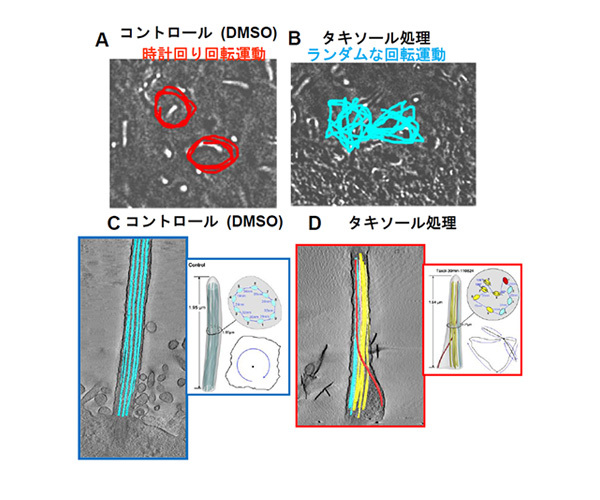
【図1】体の左右マウス胚とノード繊毛の運動と構造
(A)受精後8日目のマウス胚にあるからだの左右を決める繊毛運動。通常は時計回りに回転運動をしている。(B)マウス胚にタキソール処理を行うとノード繊毛の運動パターンが乱れる方向の定まらない回転運動と平面運動を示す。(C)電子線トモグラフィーにより得られたノード繊毛の微小管配置。微小管は繊毛内で規則正しく配置されている。(D)タキソール処理を行ったマウス胚ノード繊毛の微小管配置。微小管の配置が乱れる。膜近傍にある微小管が途中から軸糸中心へ移動している。
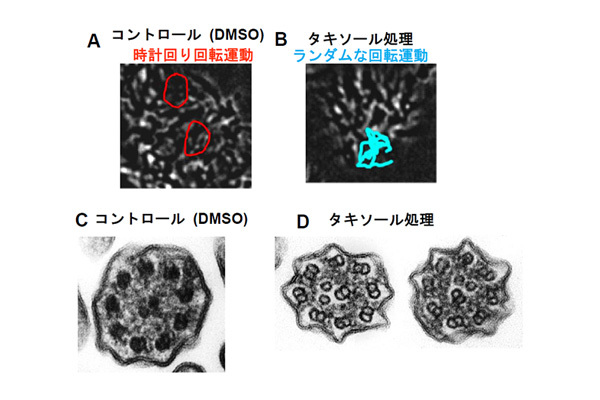
【図2】ラジアルスポークを欠損した気管繊毛は回転運動を示す。また、微小管配置が摂動に対して脆弱になる。
(A)ラジアルスポークを欠損した気管繊毛の運動。通常気管繊毛は平面運動を示すが、この変異体では時計回りの回転運動を示す。(B)タキソール処理をした気管繊毛の運動パターン。野生型気管繊毛の運動パターンはタキソール処理により変化しない。一方でラジアルスポークを欠損した気管繊毛はタキソール処理により反応し方向の定まらない回転運動になる。(C-D)ラジアルスポークを欠損した気管繊毛の断面構造。ラジアルスポークを欠損する事により気管繊毛内の微小管の配置が乱れるが、頻度は低い。一方、タキソール処理を行うと微小管の配置が乱れる頻度が増加する。