植物の生命活動を支える起動装置を発見 ―細胞膜プロトンポンプを直接活性化する基本メカニズムを解明、気孔が開く仕組みも明らかに―
植物の生命活動を支える起動装置を発見
― 細胞膜プロトンポンプを直接活性化する基本メカニズムを解明、気孔が開く仕組みも明らかに ―
発表のポイント
- 植物の細胞膜プロトンポンプを直接リン酸化して活性化するキナーゼを特定した
- C5およびC7グループのRaf型キナーゼが複合体を形成し、プロトンポンプのThr881をリン酸化する"起動装置”として機能することを発見した
- この起動原理がコケから被子植物まで保存され、4億年以上維持されてきた植物共通の「生命維持OSのコード」であることを解明した
- この起動装置に光とCO2シグナルが統合されることで朝に気孔が開く分子メカニズムを解明した
- 地球のガス交換を支える分子基盤を提示し、環境適応型作物の開発などへの応用に期待
植物は、毎朝"スイッチが入る”ことで活発な生命活動を開始します。本研究は、そのスイッチの正体を分子レベルで初めて突き止めました。本研究は、東京農工大学大学院農学研究院 生物システム科学部門の梅澤泰史教授および同大学大学院 生物システム応用科学府の高瀬緋奈乃氏(博士課程2年)を中心に、名古屋大学、山口大学、東京理科大学、宇都宮大学との共同研究として行われました。
本研究では、植物の生命活動を駆動するエンジンである細胞膜プロトンポンプ注1)を直接活性化する"起動装置”が、C5およびC7グループに属するRaf型キナーゼ注2)の複合体であることを明らかにしました。さらに、この仕組みが光と二酸化炭素(CO2)の変化に応答して働くことで、植物が毎朝気孔注3)を開くメカニズムを分子レベルで説明できることを示しました。この一連の過程は試験管内で再構成され、理論だけでなく実験的にも裏付けられました。
本研究成果は、Science(5月14日付)に掲載されました。
論文名:Raf-like protein kinase heterocomplexes directly regulate the plant plasma membrane H+-ATPase
URL:https://www.science.org/doi/10.1126/science.adx9533
背景
植物は移動することなく生きながらも、光合成によってエネルギーを獲得し、根から栄養を取り込み、呼吸し、環境の変化に適応しながら生長しています。こうした生命活動のほぼすべてを陰で支えているのが、細胞の表面に存在する細胞膜プロトンポンプです(図1)。
このタンパク質は、ATPというエネルギーを使って水素イオンを細胞外へ送り出し、細胞の内外に「見えないエネルギーの差」を生み出します。この差は、植物にとっての"動力源”として働き、栄養吸収や物質輸送を支えています(図1)。
さらに、朝、太陽の光を浴びるとプロトンポンプが活性化し、気孔が開きます。これにより植物は大気中のCO2を取り込み、光合成を開始します。この仕組みは太古の昔から地球上で毎日繰り返され、植物の物質生産を支えるとともに、CO2の吸収を通じて大気組成のバランスを保ってきました。すなわち、この分子の働きは、私たちの食料となるエネルギーを生み出すだけでなく、地球規模の炭素循環を支え、気候そのものを左右する重要な役割を担っています。
このようにプロトンポンプは、「栄養吸収」「呼吸」「生長」「環境応答」、そして「大気中CO2の調節」に至るまで、植物の生命活動を根底から支える"メインエンジン”です。しかし、この"エンジン”がどのようにして起動されるのかは、長年にわたり未解明のままでした。特に主要活性化部位の一つである881番目のスレオニン(Thr881)は近年報告されたばかりであり、これをリン酸化注4)する酵素(プロテインキナーゼ注5))の同定が、分野の重要課題となっていました。
研究体制
本研究は国立大学法人東京農工大学大学院農学研究院生物システム科学部門の梅澤泰史教授、同大学大学院生物システム応用科学府の高瀬緋奈乃氏を中心として、永野愛奈氏、神山佳明氏、山下昂太氏、片桐壮太郎氏、李揚丹氏、同大学大学院農学研究院生物生産科学部門の安達俊輔教授および連合農学研究科の川口喜暉氏、東京理科大学創域理工学部の西浜竜一教授、山内翔太助教、名古屋大学大学院理学系研究科(生命理学専攻)の木下俊則教授、林優紀助教、高橋宏二助教、山口大学大学院創成科学研究科(理学系学域)の武宮淳史教授、冨士彩紗助教、田原京佳氏、宇都宮大学バイオサイエンス教育研究センターの児玉豊教授、野口穂氏から構成される共同研究グループによって実施されました。
なお本研究は、日本学術振興会 科学研究費助成事業[JP23K27190、JP23H04192、JP22K19170]、生物系特定産業技術研究支援センター ムーンショット型農林水産研究開発事業[JPJ009237]、武田科学振興財団 生命科学研究助成[2022025571]、科学技術振興機構 先端国際共同研究推進事業(ASPIRE)[JPMJAP24A1]などの支援を受けて行われました。ASPIREによる支援は、研究終盤における集中的な推進を支え、本成果の創出に大きく貢献しました。
研究成果
◆ 1.プロトンポンプを直接起動する"アクセル”の発見
研究グループは、2021年に報告したC5グループのRaf型キナーゼ「Raf36」破壊株が小型化する現象参考文献1)に着目し、リン酸化プロテオーム解析注6)を行いました。その結果、Raf36破壊株では細胞膜プロトンポンプのThr881のリン酸化が約90%減少していることを発見しました(図2)。その後の解析により、C5キナーゼとC7キナーゼがペアを形成したときに初めて、プロトンポンプのThr881をリン酸化し、直接活性化できることを明らかにしました(図3)。さらに、この仕組みは、植物ホルモンのオーキシン注7)や光など異なる条件でも共通して働く、基本的な制御機構であることがわかりました(図2)。
また、この仕組みはコケ植物にも保存されており、植物が約4億年前に陸上へ進出して以来維持されてきた、極めて普遍的な制御機構であることが示されました(図4)。
本成果は、植物の生命活動を駆動する根幹原理、すなわち「生命を動かす基本プログラム(OSのコード)」の実体を示した発見と位置づけられます(図5)。
◆ 2.朝に気孔が開く仕組みの解明
植物が毎朝気孔を開く仕組みは、長年にわたり研究されてきました。従来は青色光による制御が中心と考えられていましたが、近年、細胞膜プロトンポンプが最大限に活性化されるためには、Thr948(主に青色光によって制御される)とThr881という2つの部位の"二重リン酸化”が必要であることが明らかになってきました。
本研究で発見したC5-C7キナーゼ複合体の一つであるHT1-CBC1は、主に赤色光シグナルに応答してThr881をリン酸化し、気孔開口を促進する役割を持つことが明らかとなりました(図6)。これにより、これまで不明であった赤色光による気孔開口の分子機構が明確になりました。
その仕組みは、図7に示したモデルとして、以下のように整理できます。
- 夜:呼吸などにより細胞内CO2濃度が高く、阻害因子(MPK4/12)がC5キナーゼ(HT1)を抑制 → 気孔は閉じる
- 日の出:青色光により"第一スイッチ”(Thr948)がオン(予備起動)
- 朝:光合成によりCO2が低下 → C5-C7キナーゼ複合体が活性化し"第二スイッチ”(Thr881)がオン
- 結果:二重スイッチが揃い、プロトンポンプが最大活性化 → 気孔が開く
さらに本研究では、この一連の経路を試験管内で再構成することに成功し、分子レベルでその正しさを実証しました(図8)。すなわち、気孔の開口は単一の光シグナルではなく、光(青色光・赤色光)とCO2濃度が統合された精密な制御システムが実装されていることが示されました。
光による気孔開口は、1898年にFrancis Darwinによって報告されて以来、長く研究されてきた古典的テーマです。本研究は、
「光・CO2・リン酸化シグナルが統合された気孔開口の統一モデル」(図7)
を確立し、130年にわたる謎に分子レベルで明確な答えを与えるものです。
研究の系譜
気孔開口の研究は、長年にわたり日本の研究者が世界をリードしてきた分野です。これまでに、
- 青色光によるプロトンポンプのリン酸化
- CO2応答因子HT1
- CBC1/2キナーゼ
などの重要な分子が、日本から世界に先駆けて発見されてきました。
本研究は、これら個別に解明されてきた要素を一つの連続した経路として統合し、さらにプロトンポンプの直接活性化機構までを実証したものです。すなわち、「日本の植物科学が積み重ねてきた知見が、一つの原理として結実した成果」といえます。
今後の展開
本成果は、植物の生命活動を駆動する基本原理の理解と、その操作を可能にするものです。これにより、
- 干ばつに強い作物(高水利用効率)の開発
- 肥料使用量を削減した持続可能な農業
- 環境ストレス耐性(高温・塩害・乾燥)の向上
- 光合成や生長の精密制御
など、幅広い応用が期待されます。
今後は、植物の生命機能を「理解する」段階から「設計・制御する」段階へと進む基盤技術としての発展が見込まれます。これは、東京農工大学が推進する地域中核・特色ある研究大学強化促進事業(J-PEAKS)の戦略目標「世界へ展開する食とエネルギーのサステイナブルイノベーション」にも合致する成果です。
用語解説
注1)細胞膜プロトンポンプ
細胞膜に存在し、ATPのエネルギーを使って水素イオン(H+)を細胞外へ排出するタンパク質。細胞の内外にエネルギーの差(電気化学ポテンシャル)を生み出し、栄養吸収や物質輸送を駆動する。
注2)Raf型キナーゼ
植物に存在するプロテインキナーゼ(タンパク質リン酸化酵素)の一群。B1~B4およびC1~C7のサブグループに分類される。本研究では、C5およびC7サブグループのキナーゼが複合体を形成し、細胞膜プロトンポンプを直接活性化することを明らかにした。
注3)気孔
葉の表面に存在する小さな孔(あな)。開閉することでCO2の取り込みや水分の蒸散を制御する。
注4)リン酸化
タンパク質にリン酸基が付加される化学反応。タンパク質の働きをオン・オフするスイッチとして機能する。
注5)プロテインキナーゼ
タンパク質にリン酸基を付加する酵素。細胞内のシグナル伝達において中心的な役割を担う。
注6)リン酸化プロテオーム解析
細胞内の多数のタンパク質について、「どのタンパク質のどの位置がリン酸化されているか」を網羅的に調べる解析手法。タンパク質の働きを制御するリン酸化の全体像を把握することで、細胞内でどのようなシグナルが働いているかを明らかにすることができる。
注7)オーキシン
植物の生長を制御するホルモンの一種。細胞の伸長を促進する働きを持つ。
参考文献1)Kamiyama, Y., Hirotani, M., Ishikawa, S., Minegishi, F., Katagiri, S., Rogan, C.J., Takahashi, F., Nomoto, M., Ishikawa, K., Kodama, Y., Tada, Y., Takezawa, D., Anderson, J.C., Peck, S.C., Shinozaki, K. and Umezawa, T. (2021) “Arabidopsis group C Raf-like protein kinases negatively regulate abscisic acid signaling and are direct substrates of SnRK2.”
Proc. Natl. Acad. Sci. USA.
118(30):e2100073118.
doi: 10.1073/pnas.2100073118. PMID: 34282011.
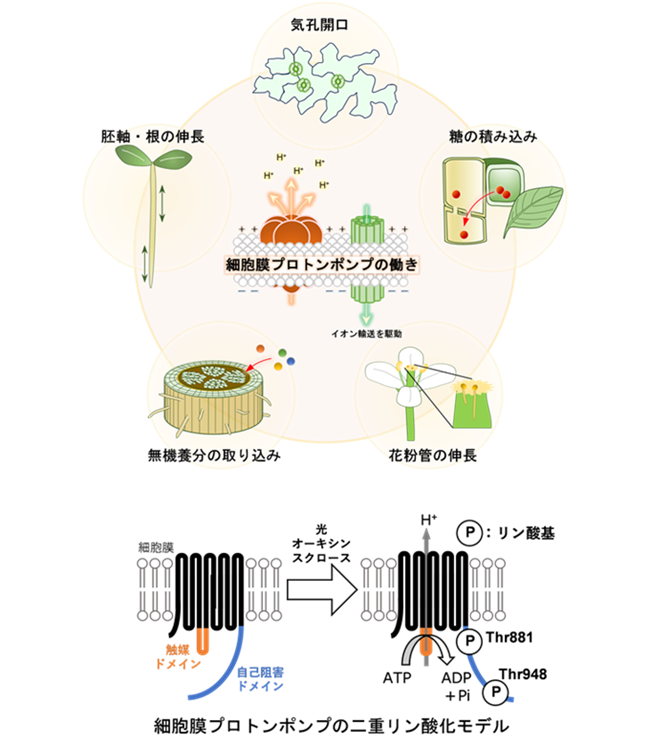
細胞膜プロトンポンプは、水素イオン(H+)を細胞外へ排出することで、さまざまなイオン輸送を駆動する膜タンパク質です。その結果、気孔開口や無機養分の取り込みなど、多様な生理応答を制御しています。細胞膜プロトンポンプには、自己阻害機構によって活性を抑える仕組みがあります。しかし、光やオーキシン、スクロースなどのシグナルを受けると、C末端領域のThr881とThr948がリン酸化されます。この二重リン酸化が、細胞膜プロトンポンプの活性化に必要であることが知られています。
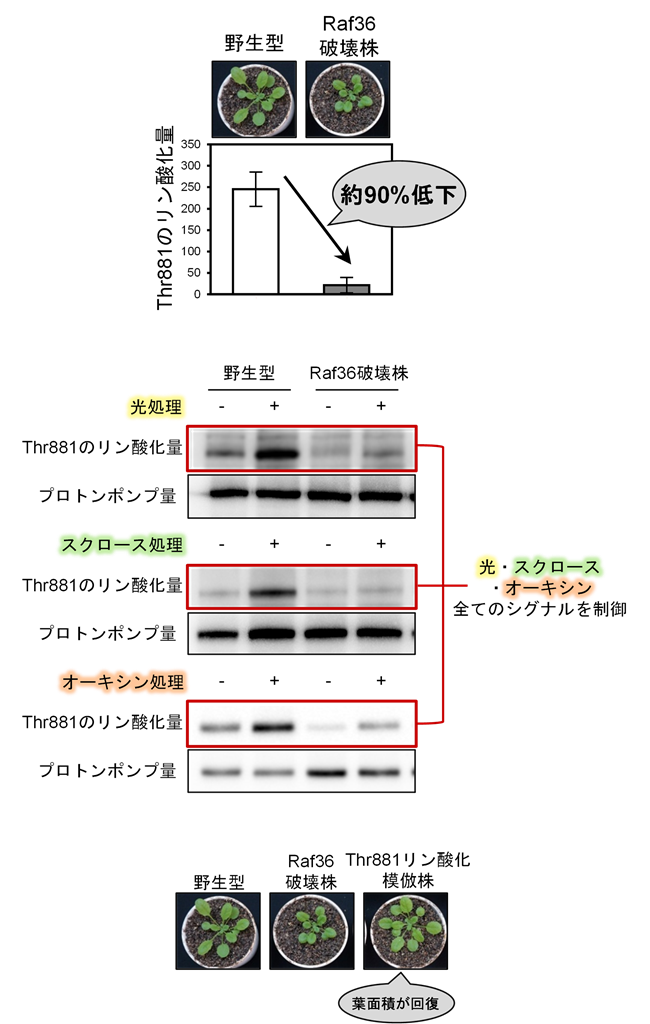
Raf36破壊株では、細胞膜プロトンポンプ活性化部位Thr881のリン酸化が約90%減少していました。さらに、Raf36による制御は、光やスクロース、オーキシンなど広範なシグナルに及ぶことがわかりました。Raf36破壊株は野生型と比較して生長が遅くなりますが、細胞膜プロトンポンプのThr881リン酸化模倣体を導入すると、この生育遅延は回復しました。これらの結果から、Raf36破壊株の生長遅延は、細胞膜プロトンポンプThr881のリン酸化低下によって引き起こされることが示されました。
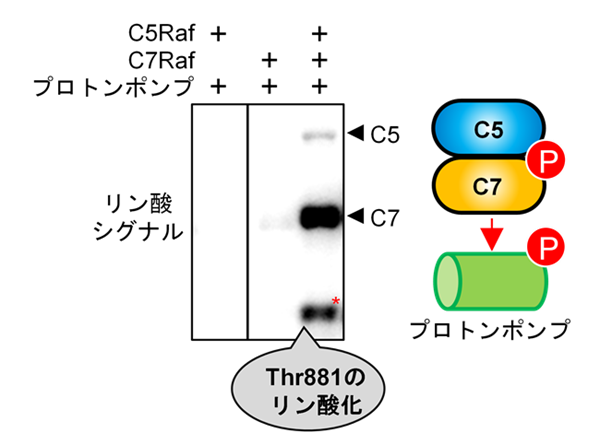
C5-RafやC7-Rafは、それぞれ単独では細胞膜プロトンポンプのThr881をリン酸化できませんでした。そこで、さまざまな実験を重ねて検討を進めた結果、C5-RafがC7-Rafをリン酸化して活性化し、その活性化したC7-Rafが細胞膜プロトンポンプThr881を直接リン酸化することを明らかにしました。この発見が、本研究のキーポイントとなりました。
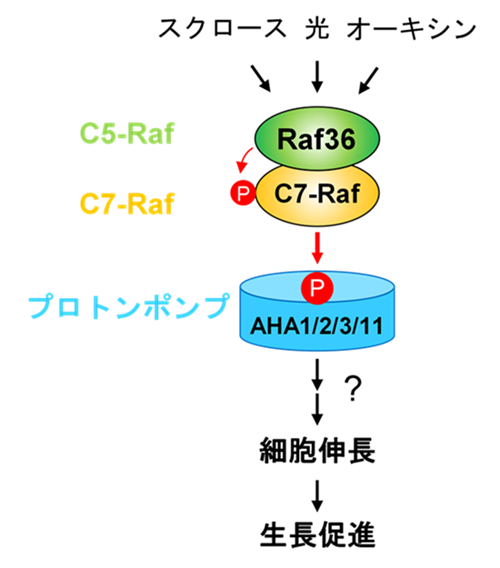
C5-RafであるRaf36は、スクロースや光、オーキシンによる細胞膜プロトンポンプの活性化を幅広く制御します。Raf36はC7-Rafを活性化し、C7-RafはThr881を直接リン酸化することでプロトンポンプを活性化します。その結果、細胞伸長や植物の生長が促進されます。
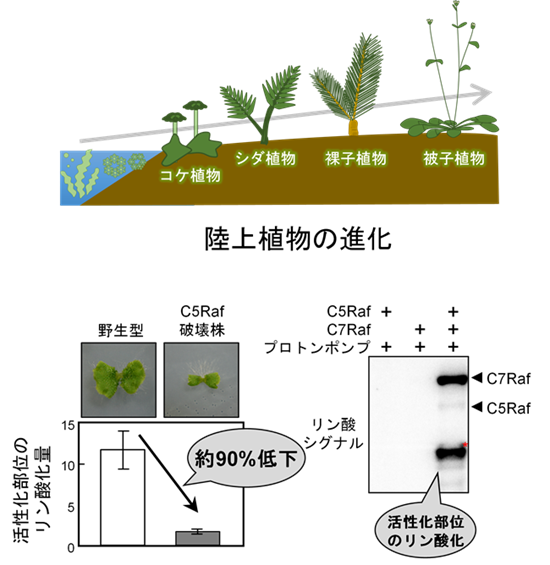
陸上植物は、その共通祖先から、維管束を持たないコケ植物の系統と、シダ植物、裸子植物、被子植物などを含む維管束植物の系統に分岐しました。コケ植物の系統でも、被子植物と同じ仕組みが働いていれば、この制御機構が陸上植物全体で広く保存されている可能性が高いと考えられます。
本研究では、ゼニゴケのC5-Raf破壊株において、野生型と比較して植物体サイズが小さくなるとともに、プロトンポンプ活性化部位のリン酸化も約90%減少していました。さらに、試験管内リン酸化試験においても、ゼニゴケのC5-C7キナーゼ複合体が活性化部位を直接リン酸化することを示しました。つまり、ゼニゴケでもシロイヌナズナと同じ仕組みが働いていることが明らかになりました。
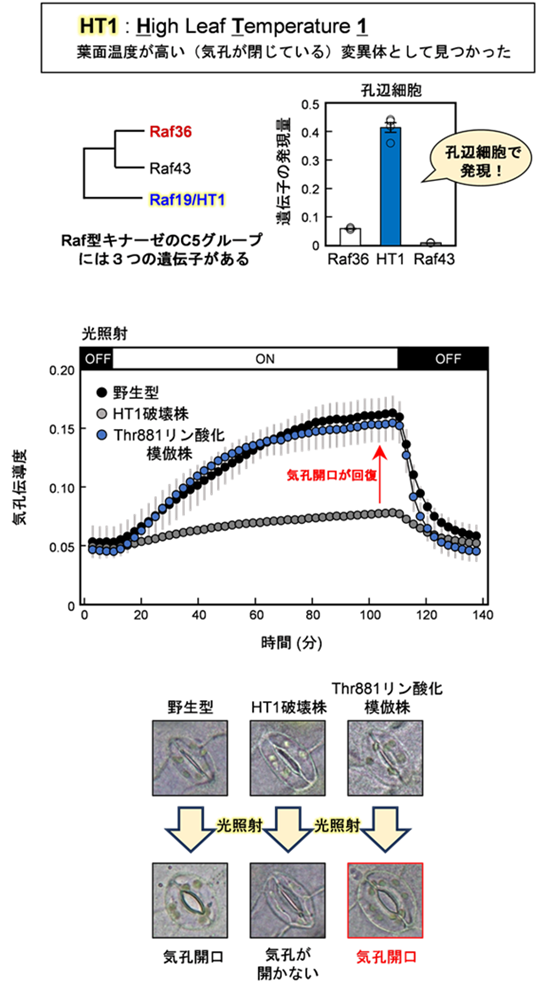
HT1は、Raf型キナーゼのC5グループに属し、孔辺細胞で多く発現しています。野生型植物では光照射によって気孔が開口しますが、HT1破壊株ではこの応答が起こりません。一方で、細胞膜プロトンポンプThr881リン酸化模倣体をHT1破壊株に導入すると、光による気孔開口応答が回復しました。これらの結果から、HT1破壊株で気孔が開かなくなる原因は、細胞膜プロトンポンプThr881のリン酸化低下によるものであることが示されました。
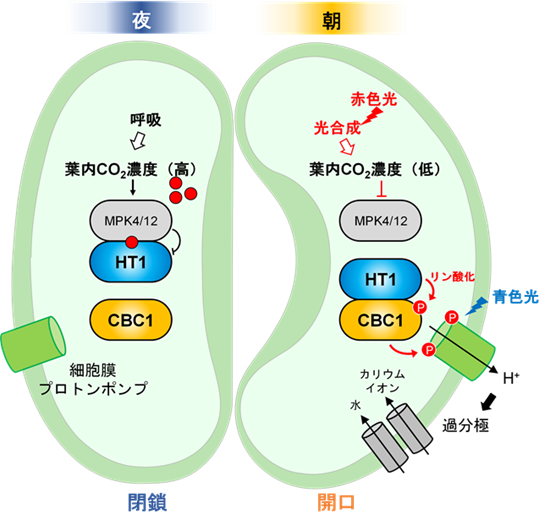
本研究で明らかとなった気孔開口のメカニズムは、次のように説明できます。
夜間は、呼吸などによって細胞内CO2濃度が高く、MPK4/12がHT1を抑制することで、気孔は閉じています。朝になると、青色光によって細胞膜プロトンポンプのThr948がリン酸化されます。さらに、太陽光が強くなるにつれて光合成が進み、細胞内CO2濃度が低下します。これによりHT1-CBC1複合体が活性化され、Thr881がリン酸化されます。この二重リン酸化が揃うことで、細胞膜プロトンポンプが最大限に活性化し、気孔が開きます。本研究では、光による気孔開口の仕組みを、光シグナルから細胞膜プロトンポンプ活性化まで一貫して説明することに、世界で初めて成功しました。
◆研究に関する問い合わせ◆
東京農工大学大学院農学研究院
生物生物システム科学部門 教授
梅澤 泰史(うめざわ たいし)
TEL/FAX:042-388-7364
E-mail:taishi(ここに@を入れてください)cc.tuat.ac.jp
◆報道に関する問い合わせ◆
東京農工大学 総務課広報室
TEL:042-367-5930
E-mail:koho2(ここに@を入れてください)cc.tuat.ac.jp
山口大学 総務部総務課広報室
TEL:083-933-5007
E-mail:sh011(ここに@を入れてください)yamaguchi-u.ac.jp
東京理科大学 経営企画部 広報課
TEL:03-5228-8107
E-mail:koho(ここに@を入れてください)admin.tus.ac.jp
宇都宮大学 広報・渉外係
TEL:028-649-5201
E-mail:kkouhou(ここに@を入れてください)a.utsunomiya-u.ac.jp
科学技術振興機構 広報課
TEL:03-5214-8404
E-mail:jstkoho(ここに@を入れてください)jst.go.jp
◆JST事業に関する問い合わせ◆
科学技術振興機構 国際部 先端国際共同研究推進室
荒川 敦史(あらかわ あつし)
TEL:03-6261-1994
E-mail:aspire(ここに@を入れてください)jst.go.jp
関連リンク(別ウィンドウで開きます)